Ánh sáng kì diệu
Trần Thanh Long
Phòng Thí nghiệm Công nghệ Snh học Phân tử, khoa Sinh học-Công nghệ Sinh học, Đại học Khoa học Tự nhiên Tp.HCM
Kỳ 4: Câu chuyện của những Prometheus
Cách mà Roger Y. Tsien, nhà khoa học thứ ba cùng chia sẻ giải Nobel hóa học 2008, đến với GFP cũng gần giống với Chalfie, ông tìm kiếm một công cụ mới giúp đỡ mình trong việc nghiên cứu. Tsien là một nhà hóa sinh đã theo đuổi việc nghiên cứu về cAMP (cyclic adenosine 3’,5’-monophosphate) thuộc hệ thống truyền tín hiệu thứ hai trong tế bào (là tập hợp các phân tử truyền tín hiệu nội bào, được tế bào giải phóng để kích hoạt các thay đổi sinh lý trong tế bào như sự phân bào, biệt hóa, di cư, sự chết tế bào…) từ năm 1988. Tsien muốn xây dựng một hệ thống cảm biến phát huỳnh quang để theo dõi hoạt động của cAMP trong tế bào, vì vậy ý tưởng theo đuổi ông là sử dụng những protein trong tự nhiên có thể gắn với cAMP. Sau rất nhiều cân nhắc từ những ứng cử viên có thể sử dụng vào thời điểm đó, Tsien và nhóm của ông quyết định sử dụng PKA (protein kinase A), một enzyme phụ thuộc cAMP và có chức năng phosphoryl hóa các protein khác. PKA gồm hai tiểu phần là tiểu phần điều hòa và tiểu phần thực hiện chức năng phosphoryl hóa. Nếu không có cAMP, tiểu phần điều hòa sẽ gắn chặt vào tiểu phần chức năng, do đó PKA sẽ không hoạt động. Ngược lại, nếu cAMP xuất hiện và gắn vào tiểu phần điều hòa, tiểu phần chức năng sẽ được giải phóng và bắt đầu chuyển nhóm phosphate từ ATP đến các protein mục tiêu. Vậy, làm cách nào Tsien có thể trực tiếp quan sát hoạt động kích hoạt PKA của cAMP. Ngay từ khi còn là sinh viên, ông đã bị cuốn hút bởi một hiện tượng sinh lý là FRET (fluorescence resonance energy transfer), hiện tượng này diễn ra khi một phân tử thuốc nhuộm bị kích thích bằng sóng ánh sáng sẽ phát ra năng lượng để kích thích một phân tử cùng loại thứ hai ở gần đó, phân tử thứ hai này sẽ phát ra ánh sáng có bước sóng nằm trong vùng khả kiến. Tsien đã ứng dụng hiện tượng này vào công trình của mình bằng cách gắn phân tử thuốc nhuộm thứ nhất vào tiểu phần điều hòa và phân tử thuốc nhuộm thứ hai vào tiểu phần còn lại. Từ đó hiện tượng FRET sẽ diễn ra khi phức hợp PKA còn nguyên vẹn. Khi có sự xuất hiện của cAMP, hai tiểu phần của PKA không còn gắn vào nhau, từ đó hai phân tử thuốc nhuộm cũng tách xa ra dẫn đến sự chấm dứt FRET. Kết quả là sự thay đổi của huỳnh quang sẽ báo hiệu sự hiện diện của cAMP. Tuy vậy, để có thể thực hiện được thử nghiệm này, Tsien cần rất nhiều protein PKA cũng như rất nhiều kĩ thuật phức tạp mà ngay cả phòng thí nghiệm của ông cũng chưa thực hiện được. Sau khi có được sự giúp đỡ từ những phòng thí nghiệm khác, Tsien cũng thành công trong việc gắn hai phân tử thuốc nhuộm là fluorescein và rhodamine lần lượt vào tiểu phần chức năng và tiểu phần điều hòa của PKA vào năm 1990. Trong những năm kế tiếp, ông tiếp tục sử dụng cảm biến cAMP dựa trên FRET do mình tạo ra để nghiên cứu một số vấn đề trong quá trình truyền tín hiệu bằng cAMP. [1]

Hình 12: Hệ thống cảm biến theo dõi cAMP dựa trên FRET của Tsien: Tiểu phần chức năng được gắn với fluorescein (Fl) (hấp thụ và phát xạ ánh sáng có bước sóng lần lược là 490 nm và 520 nm). Tiểu phần điều hòa gắn với rhodamine (Rh) (hấp thụ ánh sáng 520 nm, phát xạ ánh sáng 580 nm). Khi không có cAMP, hai tiểu phần này liên kết với nhau, khi kích thích toàn hệ thống bằng ánh sáng xanh lam 490 nm, hệ thống phát ra ánh sáng đỏ 580 nm. Khi có sự xuất hiện của cAMP, hai tiểu phần của PKA tách rời nhau, kích thích bằng ánh sáng xanh lam 490 nm sẽ làm Fl phát ra ánh sáng xanh lục còn Rh thì không phát quang. [1]
Mặc dù phát minh trên khá thành công, cách tiếp cận như trên sẽ cực kì khó khăn khi áp dụng cho những quá trình hoặc con đường chuyển hóa khác. Những ứng dụng tương tự sẽ đòi hỏi một lượng rất lớn protein mục tiêu đã được tinh sạch. Kế đến, quá trình gắn phân tử thuốc nhuộm được tiến hành in vitro trong điều kiện cực kì nghiêm ngặt để hiệu quả gắn là cao nhất cũng như các phân tử thuốc nhuộm không được ảnh hưởng đến cấu trúc và chức năng của protein mục tiêu. Sau đó là tái tinh sạch một lần nữa để thu được protein đã gắn thành công và vi tiêm vào trong tế bào sống. Tế bào mục tiêu cũng phải đủ lớn để chịu được tác động của mũi kim và có thể tự phục hồi sau đó. Cuối cùng, người làm thí nghiệm phải vô cùng kiên nhẫn và tỉ mỉ. Tsien nhận thấy rằn tất cả những khó khăn trên sẽ không còn nếu các nhà khoa học có trong tay các đoạn gene mã hóa cho các protein phát huỳnh quang. Những gene này sẽ được gắn với các gene của protein mục tiêu, sau đó, chuyển gene dung hợp này vào tế bào sống. Cả hai gene sẽ được phiên mã và dịch mã cùng nhau, cặp protein tạo thành sẽ được theo dõi thông qua sự phát huỳnh quang của protein mục tiêu. Việc chuyển gene vào tế bào cũng đơn giản hơn nhiều so với chuyển protein do mỗi tế bào chỉ cần một hoặc một vài bản sao của gene mục tiêu so với hàng tỉ phân tử protein cần chuyển vào một tế bào nếu muốn theo dõi hoạt động của protein. Cuối cùng, sau khi được chuyển gene, tế bào sẽ có nhiều thời gian để phục hồi hơn. [1]
Tất cả những lí do trên khiến Tsien vẫn chưa hài lòng với phát minh theo dõi cAMP dựa trên FRET của mình, ông lại bắt đầu tìm kiếm protein phát huỳnh quang phù hợp để cải thiện tất cả những khuyết điểm của phương pháp cũ. Ông chợt nhớ đến một số phòng thí nghiệm “đối thủ” đang sử dụng một protein phát huỳnh quang từ sứa để theo dõi ion calcium trong tế bào sống. Tuy nhiên, aequorin không phải là thứ ông cần, thay vào đó, Tsien quan tấm đến một protein tạp nhiễm khó chịu cần phải được loại bỏ cẩn thận khi tách chiết aequorin từ tự nhiên. Sau khi đào bới cơ sở dữ liệu về những công bố khoa học liên quan đến GFP, Tsien vô cùng phấn khích khi phát hiện một công trình vừa công bố của Douglas Prasher về việc dòng hóa thành công GFP cũng như khả năng lớn là chromophore của nó tự hình thành bên trong cấu trúc protein (vào thời điểm đó, đa số các protein phát huỳnh quang đã biết không thể tự phát sáng. Chúng chỉ thực hiện được chức năng phát huỳnh quang khi tiếp xúc với các phân tử hoạt hóa (cofactor) bên ngoài protein). Tsien lập tức tìm cách liên lạc với Prasher, tuy nhiên, vào lúc đó, Prasher đã quyết định ngừng nghiên cứu về GFP do khó khăn về kinh phí và chuyển hướng nghiên cứu. May mắn là Prasher đồng ý gửi cho Tsien đoạn gene GFP mà ông đã dòng hóa cũng như một số mẫu mô sứa trong trường hợp Tsien cần tách chiết các enzyme cần cho việc hình thành chromophore (vào lúc ấy, các nhà khoa học chưa biết được khả năng tự hình thành chromophore của GFP). [1]
Sau một thời gian chuẩn bị, cuối năm 1992, Tsien liên lạc lại với Prasher để nhận gene và cũng được Prasher thông báo về việc Martin Chalfie cũng có yêu cầu tương tự Tsien và đã nhận được mẫu trước ông. Một điều trùng hợp là sau đó ít lâu, Tsien và Chalfie gặp nhau ở một hội nghị khoa học. Cuộc gặp càng khiến Tsien thêm tự tin vào hướng đi của mình khi biết rằng Chalfie đã biểu hiện thành công GFP ở E. coli mà không cần sự hỗ trợ từ bất kì enzyme nào từ sứa. Tsien lập tức bắt tay vào việc, lần này ông chọn vật chủ để chuyển gene là nấm men, một sinh vật quen thuộc trong nghiên cứu di truyền, ngoài ra, lựa chọn này là để tránh trùng lặp với Chalfie do vi khuẩn thường là đối tượng đầu tiên được chọn để tiến thử nghiệm biểu hiện gene. [1]
Dĩ nhiên thử nghiệm đầu tiên của Tsien thành công do kinh nghiệm và chỉ dẫn từ Chalfie, tuy vậy, giữa các tế bào nấm men có sự khác biệt rất lớn về độ sáng chứng tỏ GFP tự nhiên thiếu ổn định khi biểu hiện ở các loài sinh vật khác. Ngoài ra, để tiếp tục theo dõi hoạt động của hai tiểu phần PKA, Tsien cần một protein phát huỳnh quang khác GFP. Tìm kiếm một protein mới từ tự nhiên chắc chắn khó hơn rất nhiều với việc cải thiện những gì đã có. Tsien quyết định biến đổi GFP để thu được những dẫn xuất ổn định hơn cũng như phát sáng nhiều loại màu sắc hơn. Ban đầu, do tyrosin ở vị trí 66 là một trong ba amino acid cấu tạo nên chromophore, Tsien thử thay thế tyrosine bằng tryptophan do amino acid này mang vòng thơm, cấu trúc dễ hấp thụ ánh sáng và phát huỳnh quang. Tuy nhiên, thử nghiệm này thất bại khi GFP đột biến hoàn toàn mất khả năng phát sáng. Điều này khiến nhóm của Tsien quyết định đi theo cách mà tự nhiên đã làm từ hàng tỉ năm nay để có những đặc tính phù hợp nhất, đột biến và chọn lọc. Nhóm của ông gây đột biến ngẫu nhiên trên gene của GFP, sau đó chuyển vào nấm men và chọn ra những dòng nấm men phát ánh sáng phù hợp, sau đó quay ngược lại giải trình tự gene để xem đột biến nào gây ra đặc điểm này. Sau nhiều thử nghiệm, nhóm của Tsien đã tạo ra hai loại đột biến đầu tiên có thể phát sáng xanh lam là BFP (blue fluorescent protein, histidine thay thế tyrosin ở vị trí 66) và CFP (cyan fluorescent protein, thay thế tyrosine ở vị trí 66 bằng tryptophan. Sự thay đổi này giống với dự đoán của Tsien trước đây, tuy nhiên lần thử nghiệm này thành công là do ngoài thay thế tyrosine 66, dẫn xuất này còn có một số đột biến để tạo không gian đủ lớn trong protein để chứa đủ một amino acid lớn như tryptophan). CFP càng hữu dụng hơn khi có đặc tính không bị phai màu. [1]
Về cơ bản, cùng với GFP, Tsien đã có thêm 2 màu nữa đủ để lặp lại thử nghiệm cảm biến cAMP bằng PKA. Tuy nhiên, do CFP tạo ra một màu sắc trung gian giữa xanh lam và xanh lục, việc kết hợp nó với GFP (ánh sáng xanh lá) hoặc BFP (ánh sáng xanh lam) để gắn lên các tiểu phần của PKA sẽ không phù hợp do khó nhận biết sự thay đổi về màu sắc huỳnh quang. Vì vậy, kết hợp GFP với BFP là phương án khả dĩ. BFP sẽ được kích thích bằng tia UV, ánh sáng xanh lam từ BFP sẽ được GFP hấp thụ và phát ra ánh sáng xanh lục khi hai protein này nằm gần nhau trong trường hợp không có cAMP. Ngược lại, khi có cAMP, hai tiểu phần của PKA tách nhau ra dẫn đến sự chia tách GFP và CFP, kích thích bằng tia UV sẽ chỉ làm BFP phát sáng xanh lam. Tuy nhiên, GFP ngoài hấp thụ ánh sáng xanh lam còn có thể hấp thụ tia UV và thậm chí, khả năng hấp thụ UV còn mạnh hơn nhiều so với ánh sáng xanh lam, sau đó tự phát ra ánh sáng xanh lục mà không cần BFP. Vì vây, dù trong trường hợp nào, GFP cũng phát sáng và không thể chỉ ra sự xuất hiện của cAMP hay không. [1]
Vấn đề trên thôi thúc Tsien phải cải thiện được GFP hoặc tạo ra được một protein phát huỳnh quang mới. Tiếp tục gây đột biến, Tsien đã tìm được thêm hai loại protein mới: GFP cải tiến (serine 65 thay bằng threonine), không còn hấp thụ tia UV, đồng thời độ hấp thụ ánh sáng xanh lam tăng lên 5-6 lần. Sự thay đổi từ serine thành threonine chỉ khác nhau ở một nhóm methyl, nhưng như vậy cũng đủ để thay đổi mạng lưới liên kết hydro giúp ổn định trạng thái trung hòa của chromophore, từ đó, chromophore đột biến chỉ tồn tại ở dạng anion nên chỉ hấp thụ ánh sáng xanh lam (xem lại phần 1). Kết quả thứ hai là YFP (yellow fluorescent protein). Trước đó, Jim Remington, một nhà khoa học ở đại học Oregon, sau khi nghiên cứu cấu trúc GFP đã chú ý tới một hốc trong lòng GFP gần với chromophore. Hốc này đủ lớn để chứa một vòng thơm, từ đó vòng thơm mới xuất hiện này có thể tương tác không cộng hóa trị với vòng thơm của chromophore thông qua các liên kết π trên cả hai vòng. Tương tác mới hình thành này có thể thay đổi bước sóng hấp thụ và phát xạ của chromophore. Dựa trên dự đoán trên, nhóm của Tsien đã thay thế threonine 203 bằng một vài amino acid có vòng thơm, và các đột biến làm tăng bước sóng hấp thụ lẫn phát xạ cực đại của GFP thêm 20 nm, phát ra ánh sáng vàng. Với các dẫn xuất mới này, Tsien đã có thể thoải mái ứng dụng vào việc theo dõi cAMP của mình. [1]

Hình 13: Bốn dẫn xuất của GFP tự nhiên được biểu hiện trên vi khuẩn. Theo chiều kim đồng hồ: protein phát sáng xanh lam; protein phát sáng xanh cyan; GFP cải tiến; protein phát sáng vàng. [1]
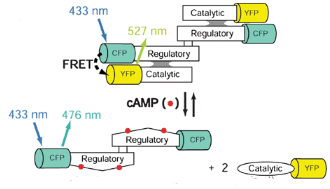
Hình 14: Hệ thống cảm biến theo dõi cAMP dựa trên FRET cải tiến: các phân tử thuốc nhuộm đã được thay thế bằng protein phát huỳnh quang là CFP và YFP. [1]
Ông cũng là người tìm ra cơ chế chromophore tự hình thành trong GFP với điều kiện duy nhất là oxi. Nhận định rằng ba amino acid tạo nên chromophore được bảo vệ quá kĩ lưỡng từ các phiến β và xoắn α xung quanh, không enzyme nào có thể tiến vào trong để xúc tác sự hình thành chromophore, vì vậy đây là một phản ứng không cần enzyme. Kế đến, việc hình thành chromophore bao gồm sự dehydro hóa, hai nguyên tử hidro được giải phóng cần phải có phân tử tiếp nhận và đó có thể là khí oxi. Do oxi hiện diện trong tất cả các loài sinh vật trừ những loài kị khí bặt buộc, Tsien chỉ cần chuyển gene GFP vào vi khuẩn trong điều kiện kị khí chặt chẽ, và đúng như dự đoán, những tế bào vi khuẩn không phát sáng. Sau khi vi khuẩn tiếp xúc với không khí, ánh sáng xanh lục dần hình thành sau vài giờ.
Chương cuối cùng trong quá trình phát triển các protein phát huỳnh quang bắt đầu với phát hiện về protein phát huỳnh quang tương tự GFP tìm thấy ở san hô. Matz và cộng sự đã phân lập được gene mã hóa cho protein này ở một loài san hô trong thủy cung ở Moscow. Tsien nhanh chóng bắt tay vào tìm hiểu protein mới này và bất ngờ phát hiện chromophore của nó gần như giống hệt của GFP. Quá trình hình thành của hai loại chromophore cũng gần như giống hệt nhau, chỉ khác là chromophore mới này được dehydro hóa một lần nữa để tạo nên một nối đôi mới thứ hai. Sự thay đổi này làm cho protein này phát huỳnh quang đỏ và được đặt tên là DsRed (Discosoma sp. red fluorescent protein). Nhiều nhà khoa học sau đó đã cải tiến DsRed để ứng dụng như GFP. Do DsRed trong tự nhiên là một protein bậc 4 gồm 4 tiểu phần, nó có cấu trúc phức tạp hơn GFP và rất khó để biểu hiện gene của DsRed với gene của protein mục tiêu như ở GFP (do gene dung hợp phải được biểu hiện 4 lần mới tạo được 1 phân tử DsRed hoàn chỉnh). Nhưng nỗ lực của nhiều nhóm nghiên cứu đã tạo được DsRed cải tiến chỉ có một tiểu phần (RFP-red fluorescent protein). Các dẫn xuất của RFP đã bao phủ toàn bộ phần còn lại của phổ ánh sáng khả kiến. [2]
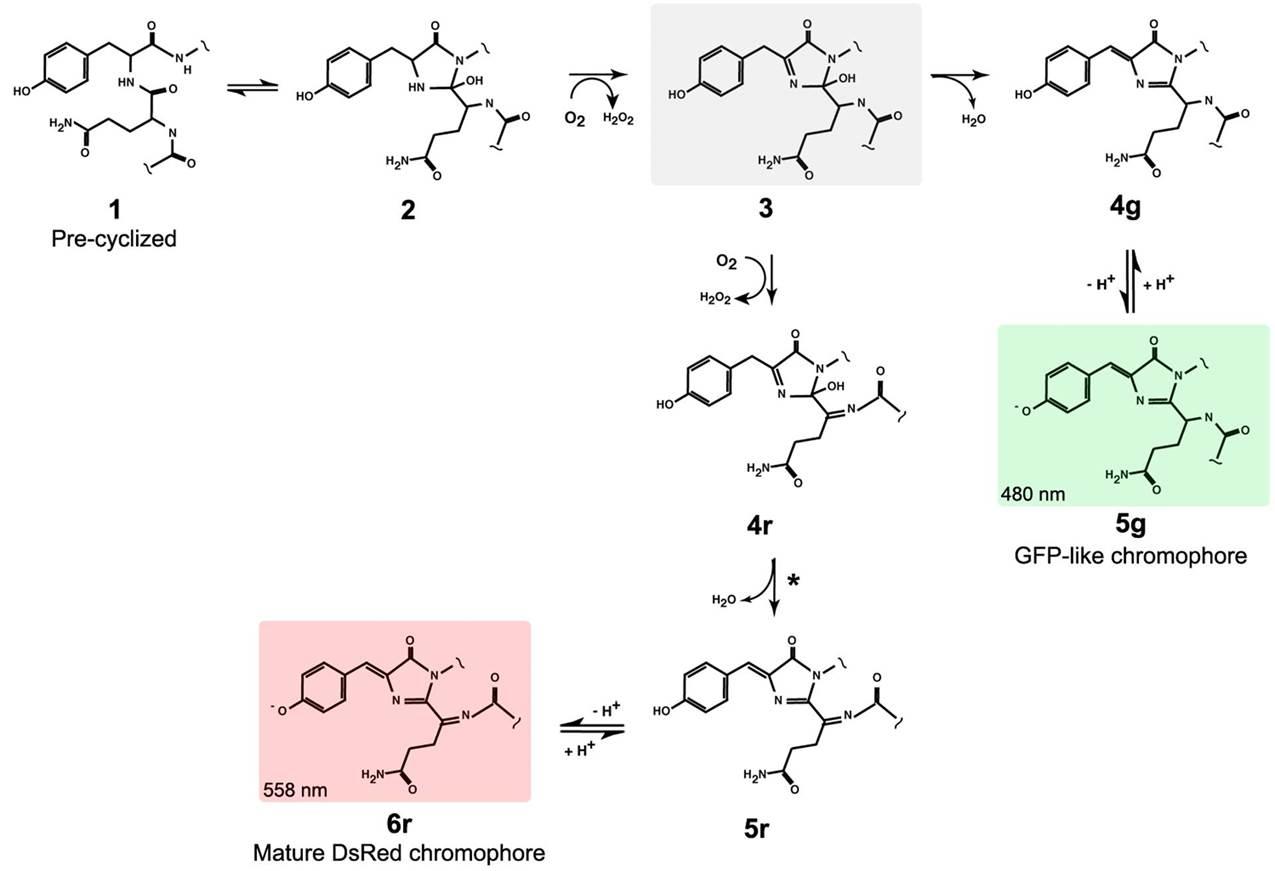
Hình 15: So sánh quá trình hình thành chromophore của GFP và DsRed. Sự hình thành chromophore của DsRed chỉ khác biệt với GFP ở bước số 3 khi chromophore của DsRed trải qua thêm một bước dehydro hóa, kết quả là chromophore của DsRed có thêm một nối đôi so với chromophore GFP, từ đó làm cho cấu trúc này có thể hấp thụ và phát xạ photon với bước sóng khác xa GFP. [2]
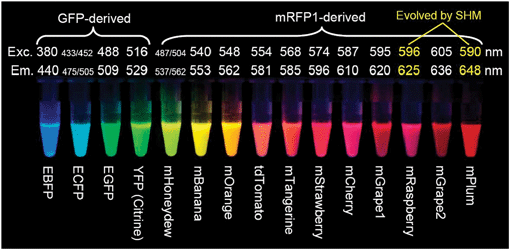
Hình 16: Các biến thể của GFP và mRFP1 (là DsRed cải tiến khi chỉ gồm một tiểu phần duy nhất). Exc: bước sóng kích thích của từng dẫn xuất. Em: bước sóng phát xạ của các dẫn xuất. [1]
Nổi tiếng khi là một trong những người tiên phong mở rộng tiềm năng của protein phát huỳnh quang, Tsien thường được hỏi về những ứng dụng của các protein phát huỳnh quang này. Ví dụ ưa thích của ông là việc theo dõi virus HIV bằng GFP. Gắn gene GFP vào bộ gene HIV giúp các nhà khoa học bất ngờ nhận ra rằng một khi đã xâm nhập vào tế bào đầu tiên, HIV sẽ xâm nhiễm sang các tế bào khác thông qua các cầu nối đặc biệt hình thành giữa các tế bào. Cầu nối này do virus kích thích các tế bào tạo thành. Qua cầu nối, virus sẽ xâm nhiễm trực tiếp từ tế bào bệnh sang tế bào lành mà không cần tiếp xúc với chất nền ngoại bào. Vì vậy, các nỗ lực tìm kiếm vaccine để ức chế HIV là vô ích (do các nhà khoa học muốn tạo ra các kháng thể úc chế được HIV để làm vaccine, tuy nhiên, kháng thể chỉ có thể tác động đến các tác nhân ngoại bào). [3]
Một tin đáng buồn là Tsien không thể tiếp tục đóng góp cho khoa học khi ông đã qua đời vào ngày 24 tháng 8 năm 2016 vì lí do mà đến nay vẫn chưa được làm rõ. Những thành tựu của ông vẫn sẽ tiếp tục hỗ trợ giới khoa học trong nhiều khám phá trong tương lai. [4]
Câu chuyện về ba nhà khoa học đoạt giải Nobel 2008 là câu chuyện đầy cảm hứng: Osamu đầy kiên nhẫn và quyết tâm thỏa mãn trí tò mò về sự phát quang sinh học trong hàng chục năm trời, Chalfie và Tsien với niềm đam mê đề tài nghiên cứu của riêng mình đã tìm kiếm, cải tiến và ứng dụng tài tình khám phá của các nhà khoa học khác. Không những vậy, những đóng góp của hai ông đã giúp giới khoa học thấy nhiều hơn và tiến xa hơn. Họ mang ánh sáng từ tự nhiên về để soi sáng chính họ và đồng thời là cho cả thế giới.
Tài liệu tham khảo:
|
[1] |
R. Y. Tsien, "Constructing and exploiting the fluorescent protein paintbox," in The Nobel prize in chemistry 2008, 2008. |
|
[2] |
"Biochemistry lab – Deciphering chromophore biogenesis," Dynamic DNA, [Online]. Available: http://www.dynamicdna.org/biochemistry-lab-course/. |
|
[3] |
"Roger Y. Tsien," Wikipedia, [Online]. Available: https://en.wikipedia.org/wiki/Roger_Y._Tsien. |
|
[4] |
"2008 nobel Laureates - The Nobel Prize in Chemistry 2008," 2008. [Online]. Available: https://www.nobelprize.org/nobel_prizes/nobelguide_che.pdf. |
|
[5] |
"The Nobel Prize in Chemistry 2008 - Illustrated presentation," Nobelprize.org, 2008. [Online]. Available: https://www.nobelprize.org/nobel_prizes/chemistry/laureates/2008/illpres.html. |
|
[6] |
O. Shimomura, "Discovery of green fluorescent protein, GFP," in The Nobel prize in chemistry, Stockholm, 2008. |

