Nguyễn V. Cường1
1Khoa Di truyền chọn giống cây trồng, Đại học Cornell, Bang New York, Hoa Kỳ.
22Department of Cancer Biology, the University of Texas M.D. Anderson Cancer Center, Houston, TX, USA 77054.
Biên tập viên: Hương Hà, Stanford University, Stanford, California, USA
* Mọi thắc mắc về bài viết xin liên hệ email cvn6@cornell.edu
Tóm tắt: Ở cây trồng, các tính trạng nông học quan trọng như năng suất thường là các tính trạng số lượng chịu ảnh hưởng của nhiều gene khác nhau và ảnh hưởng của môi trường. Đối với các tính trạng này việc xác định các locus điều khiển tính trạng (Quantitative Trait Locus-QTL- mapping) được dựa trên việc phân tích kiểu gene và kiểu hình ở các quần thể phân li và áp dụng các công cụ thống kê nhất định.Bài phân tích này giới thiệu sơ qua một phương pháp QTL mapping cơ bản thông qua một nghiên cứu về năng suất lúa.
Abstract: In crop plants, important agronomical traits like yield are quantitative traits influenced by many genes and by the environment.Theidentification of quantitative trait loci (QTL mapping) for these traits is based on analyzing genotypes and phenotypes in segregating populations and applying appropriate statistical tests. This article briefly introduces a basic QTL mapping method applied in a study of rice yield.
Từ khoá: Phân tích QTL | chỉ thị phân tử | tách dòng gene | năng suất.
Giới thiệu
Sinh học thực vật là một ngành khoa học dựa nhiều vào các phương pháp và kỹ thuật và việc nắm chắc các phương pháp và kỹ thuật này sẽ giúp nghiên cứu được nhiều vấn đề cụ thể khác nhau.Trong bài tóm lược này tôi không định tổng hợp các kết quả nghiên cứu mà muốn mượn một bài báo xuất sắc (1) để giới thiệu sơ qua một phương pháp phân tích tính trạng số lượng ở thực vật.
Ashihaki et al (1) khảo sát sự khác biệt về năng suất ở hai giống lúa và bằng việc phân tích kiểu hình và kiểu gene của quần thể tạo ra từ hai giống này đã tìm ra được chính xác những gene liên quan đến năng suất, một trong những tính trạng quan trọng nhất ở cây trồng.
Năng suất là một tính trạng số lượng phức tạp.Về cơ bản nó là tổng hợp của nhiều tính trạng khác nhau. Năng suất có hệ số di truyền thấp, ảnh hưởng lớn bởi các yếu tố môi trường. Năng suất được tính là số tấn/hecta. Ở cây ngũ cốc hạt nhỏ như lúa, năng suất có thể được tính là tích số giữa số lượng hạt lúa trên cây và trọng lượng hạt. Năng suất là một đặc tính mang tính quần thể vì trong thực tế sản xuất tính trạng này luôn được so sánh trên một diện tích sản xuất với hàng nghìn cây chứ không chỉ trên một cá thể đơn lẻ, song nghiên cứu trên quy mô nhỏ sẽ là bước khởi đầu để khảo sát đại trà trên diện tích lớn hơn. Để tăng năng suất, các nghiên cứu thường tập trung vào tăng số lượng hạt lúa trên một bông và số bông trên cây. Đây đều là các đại lượng có thể đo đếm được một cách chính xác, một điều kiện hết sức quan trọng trong phân tích di truyền số lượng, bởi nếu không có được phương pháp xác định chính xác kiểu hình tính trạng cần nghiên cứu thì dù có áp dụng các kĩ thuật phân tích phân tử và thống kê sau này, kết quả tìm ra cũng sẽ không phản ánh đúng bản chất di truyền của tính trạng.
Phương pháp xác định locus tính trạng số lượng
Như đề cập ở trên, năng suất là một tính trạng số lượng, sự khác biệt về năng suất không phải do sự phân li của một hoặc hai gene mà là do sự phân li của rất nhiều gene, với ảnh hưởng của mỗi gene là nhỏ. Tính trạng số lượng không phân li thành các nhóm cụ thể do đó ta không thể đơn giản chỉ sử dụng các nguyên lí di truyền Mendel để nghiên cứu. Thay vào đó, giải pháp cho nghiên cứu các tính trạng này là xác định vị trí các locus của tính trạng số lượng dựa trên sự liên kết với marker phân tử (QTL mapping). Ở thực vật, có hai phương pháp chính trong QTL mapping: 1) Sử dụng các quần thể tạo ra từ các phép lai được kiểm soát (ví dụ như quần thể F2, quần thể lai trở lại...) 2) Sử dụng các quần thể tự nhiên, giao phấn tự do không kiểm soát (phương pháp này còn được gọi là lập bản đồ kết hợp - association mapping). Phương pháp 1) phù hợp cho các loài có vòng đời ngắn, dễ lai tạo và tạo nhiều hạt ở thế hệ sau, đồng thời có phương thức tính toán đơn giản hơn. Phương pháp 2) có lợi thế là không phải tạo ra quần thể phân li, phù hợp cho các loài giao phấn, vòng đời dài và có thể phân tích được nhiều allele cùng một lúc. Tuy vậy phương pháp 2) có nhiều yếu tố gây nhiễu do sự giao phấn và chọn lọc không được kiểm soát và phép thống kê phức tạp hơn. Ở đây, Ashihaki et al (1) sử dụng phương pháp 1) với quần thể được tạo ra từ phép lai hai bố mẹ.Việc phân tích thống kê kiểu gene và kiểu hình của các cá thể ở quần thể phân li sẽ giúp tìm ra các vùng trên nhiễm sắc thể có thể có liên quan với tính trạng cần nghiên cứu. Kết quả thống kê cũng cho biết được sự ảnh hưởng mạnh hay yếu của vùng nhiễm sắc thể đó với tính trạng để có các nghiên cứu tiếp theo. Các yêu cầu cho QTL mapping từ các quần thể lai có kiểm soát được liệt kê ở bảng bên. Hiện nay, các kỹ thuật xác định kiểu gene và giải trình tự nhanh chóng đã giúp cho việc lập bản đồ phân tử và xác định kiểu gene các cá thể dễ dàng hơn.
Các yêu cầu cho QTL mapping từ các quần thể lai có kiểm soát
Có các marker phân tử (trung hòa, đồng trội)
Có bản đồ di truyền liên kết phân tử
Các quần thể có sự “mất cân bằng liên kết” (linkage disequilibrium - LD). LD có thể được hiểu một cách đơn giản là khi biết kiểu gene của một locus có thể suy ra được kiểu gene của locus khác - thường là do nằm cạnh nhau trên nhiễm sắc thể
Có phương pháp đo lường kiểu hình tính trạng tin cậy
Có phương pháp thống kê /phần mềm để xác định sự liên kết giữa marker và tính trạng
Ashikari et al (1) chọn hai bố mẹ là hai giống lúa khác nhau về năng suất, một giống thuộc dòng lúa ôn đới japonica có năng suất không cao nhưng chất lượng tốt, tạo ra loại gạo Koshihikari (gạo được dùng trong sushi) và một giống lúa nhiệt đới indica có năng suất cao hơn nhưng chất lượng không cao bằng, tạo ra loại gạo Habataki. Điểm cần chú ý ở đây là ngoài việc các tác giả chọn hai giống có năng suất khác nhau nhưng cũng đồng thời có sự khác biệt về nguồn gốc và khu vực phân bố nhất định (japonica và indica) để có nhiều đa hình về trình tự DNA giữa hai bố mẹ, tạo thuận lợi cho thiết kế marker phân tử cho phân tích sau này.Thêm vào đó, hai giống bố mẹ được chọn là các giống lúa thương phẩm nên kết quả nghiên cứu có thể áp dụng cho sản xuất dễ dàng hơn.
Song song với năng suất (cụ thể là số lượng hạt trên bông lúa và số bông trên cây), các tác giả cũng nghiên cứu một tính trạng nữa là chiều cao cây vì hai giống bố mẹ có chiều cao cây khác nhau. Trong lịch sử chọn giống lúa, giống thấp cây IR8 (Viện nghiên cứu lúa thế giới (IRRI)) giúp cây đứng thẳng, không bị nghiêng đổ. Đặc điểm này giúp cây phát triển và quang hợp tốt hơn khi được chăm bón tốt, giúp tăng năng suất cây trồng. Giống thấp cây này có thể coi là cơ sở cho “cuộc cách mạng xanh” ở châu Á những năm 60 của thế kỉ 20.
Từ hai giống bố mẹ trên, việc tiếp theo là tạo ra các quần thể phân li từ hai bố mẹ. Quần thể phân li có thể ở thế hệ F2 hay thế hệ lai trở lại. Ở đây Ashikari et al (1) đã tạo ra một quần thể khá đặc biệt để bắt đầu nghiên cứu là quần thể backcross inbred lines (BIL)- lai trở lại cận giao tái tổ hợp. Quần thể này được tạo ra bằng cách thế hệ F1 được lai trở lại với một trong hai bố mẹ, ở đây là giống japonica Koshihikari, thay vì tự thụ phấn. Sau đó sử dụng phương pháp một hạt một thế hệ (single seed descent) để chọn ra các dòng ở các thế hệ tiếp theo, đến thế hệ F7 khi gần như hệ gene của các dòng đã ở trạng thái đồng hợp tử. Vì sao tác giả lại tạo ra quần thể này? Như đã đề cập ở trên, tính trạng năng suất có ảnh hưởng lớn từ môi trường, và tính trạng này thường được xác định thông qua giá trị trung bình trên nhiều cá thể chứ không phải một cá thể. Việc tạo ra các dòng đồng hợp tử như vậy giúp họ có thể lặp lại được việc đo năng suất ở thế hệ tiếp theo (trồng nhiều cây hơn) để có được số liệu đáng tin cậy về năng suất mà vẫn giữ nguyên được kiểu gene của cây cần đánh giá. Quá trình tạo ra các dòng BIL này cũng giúp họ tạo được các dòng gần đẳng gene (nearly isogenic lines) để tiện lợi cho việc cô lập chính xác gene sau này. Hơn thế nữa, do đây là các dòng đồng hợp tử nên có thể trồng để nghiên cứu nhiều kiểu hình khác nhau nhưng chỉ cần xác định kiểu gene một lần, thuận lợi cho việc hợp tác nghiên cứu về sau.
Các tác giả sử dụng 96 dòng BIL để tiến hành phân tích QTL. Yêu cầu tiếp theo là phải có được một bộ các marker phân tử trên toàn hệ gene và lập bản đồ liên kết các marker đó trên quần thể đang phân tích.Việc lập bản đồ liên kết về bản chất là dựa trên hiện tượng phân li, liên kết gene và tái tổ hợp ở quần thể phân li đang phân tích để xác định vị trí các marker trên nhiễm sắc thể. Trước hết ta cần có các marker phân tử, thể hiện sự đa hình về trình tự DNA giữa hai hệ gene bố và mẹ. Các đa hình DNA thường được dùng cho phân tích bản đồ gene gồm: 1) Sai khác về một nucleotide (Single Nucleotide Polymorphisms – SNPs), 2) Đa hình do mất hoặc chèn thêm nucleotide (deletion/insertion), 3) Đa hình do thay đổi số bản sao của các trình tự lặp lại liên tiếp (như microsatellites...). Đây là đều là các loại đa hình có thể xác định được dựa trên phương pháp PCR và cắt enzyme giới hạn thông dụng trong các phòng thí nghiệm. Như đã đề cập ở trên, hai dòng lúa bố mẹ trong nghiên cứu này thuộc hai nhóm japonica và indica. Hai nhóm này có sự cách li trong lịch sử và nghiên cứu cho thấy có nhiều đa hình DNA.Thêm vào đó, trình tự lúa nhóm japonica đã được giải mã tại thời điểm tiến hành nghiên cứu nên việc phát triển bộ marker phân tử giữa hai nhóm giống này trên toàn hệ genome là điều có thể làm được.
Ashikari et al (1) chọn phân tích 200 marker trải đều trên 12 nhiễm sắc thể của lúa. Sau khi đã có các marker và xác định được kiểu gene của từng cây trong quần thể phân tích, họ dùng các công cụ tính toán để có thể lập nhóm liên kết và tính khoảng cách di truyền giữa các marker. Về cơ bản, quá trình này gồm hai phép tính: 1) xác định sự phân li độc lập hay sự liên kết giữa 2 marker, 2) nếu có sự liên kết thì xác định trật tự và khoảng cách của các marker dựa trên tần số trao đổi chéo. Với số lượng marker lớn lên tới hàng trăm, thậm chí hàng nghìn việc tính toán bằng tay là không thể. Hiện nay có nhiều phần mềm giúp ta việc tính toán này, có thể kể đến các chương trình thông dụng như Mapmaker, Joinmap hay R/qtl.
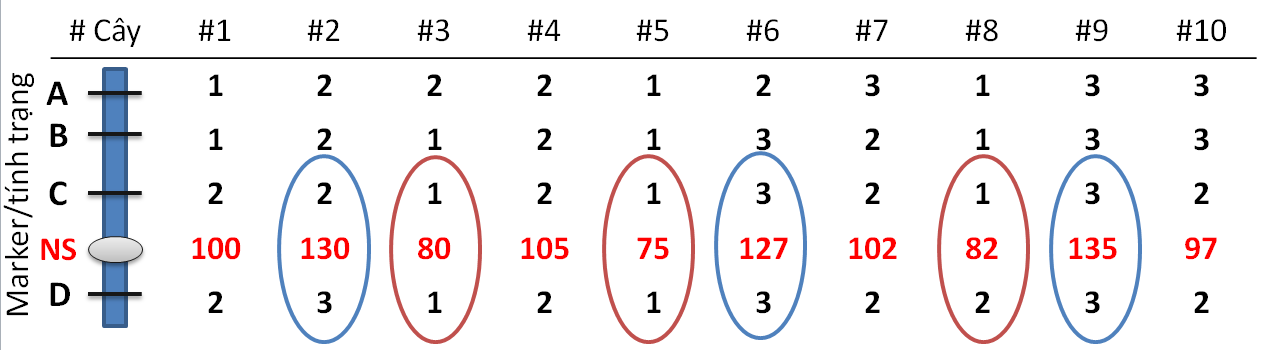 Hình 1. Ví dụ về sự liên kết giữa marker với tính trạng (NS-Năng suất). Cây đồng hợp tử về allele của mẹ được kí hiệu 1, cây đồng hợp tử về allele của bố, 3, cây dị hợp tử, 2. A, B, C, D là các marker. Ở 2 marker C và D, các cây mang allele của bố, 3, có năng suât cao hơn các cây có mang allele của mẹ, 1, thể hiện có sự liên kết giữa gene và tính trạng. Sự sai khác về kiểu hình này có thể kiểm tra bằng phép tính t -student hay ANOVA.
Hình 1. Ví dụ về sự liên kết giữa marker với tính trạng (NS-Năng suất). Cây đồng hợp tử về allele của mẹ được kí hiệu 1, cây đồng hợp tử về allele của bố, 3, cây dị hợp tử, 2. A, B, C, D là các marker. Ở 2 marker C và D, các cây mang allele của bố, 3, có năng suât cao hơn các cây có mang allele của mẹ, 1, thể hiện có sự liên kết giữa gene và tính trạng. Sự sai khác về kiểu hình này có thể kiểm tra bằng phép tính t -student hay ANOVA.
Sau khi đã xác định được kiểu gene của từng cá thể trong quần thể phân tích và lập được bản đồ gene liên kết, việc tiếp theo là xác định giá trị kiểu hình tính trạng nghiên cứu trên từng cá thể. Khi đã có số liệu kiểu gene và kiểu hình, câu hỏi đặt ra lúc này là liệu có sự liên quan giữa từng vị trí marker trên bản đồ với kiểu hình tính trạng nghiên cứu. Đối với các quần thể phân li dựa trên việc lai 2 bố mẹ như trong bài báo đang phân tích thì, về bản chất, đó là việc kiểm tra xem liệu có sự sai khác một cách có ý nghĩa thống kê về kiểu hình của các cá thể mang allele từ bố so với các cá thể mang allele từ mẹ ở một vị trí marker nhất định (Hình 1). Có nhiều phương pháp để kiểm tra việc này nhưng phương pháp cơ bản nhất là kiểm tra bằng phép toán t-student hoặc phân tích phương sai (ANOVA).
Trên thực tế phương pháp phân tích QTL phổ biến được dùng là phương pháp interval mapping (dò tìm theo đoạn) (2), ước lượng vị trí QTL giữa hai marker và sử dụng phép thống kê là giá trị ODDs (viết dưới dạng logarithm of ODDs - LOD). Giá trị ODDs là tỉ số giữa xác suất sự kiện 1 (H1) trên xác suất sự kiện 2 (H2). Ví dụ: Ta tham gia vào trò chơi tung đồng xu và đặt cược sự xuất hiện mặt sấp, người chủ trò tung 10 lần liên tiếp nhưng đều là mặt ngửa! Liệu có sự gian lận ở đây? Xác suất để một “đồng xu” có hai mặt đều là mặt ngửa (người chủ trò đã gian lận) tung 10 lần liên tiếp là mặt ngửa (H1) là (1)10=1. Xác suất để một đồng xu có hai mặt (sấp và ngửa, tức là đồng xu thật) tung 10 lần liên tiếp là mặt ngửa (H2) là (0.5)10 = 10-3. Giá trị ODDs ở đây sẽ là H1/H2 = 1000 và LOD là 3, có nghĩa là khả năng người chủ trò sử dụng đồng xu có hai mặt đều là mặt ngửa (gian lận) cao hơn gấp 1000 lần khả năng sử dụng đồng xu có hai mặt sấp và ngửa (đồng xu thật).
Một điểm cần chú ý trong phân tích QTL đó là số lượng các phép kiểm tra như trên. Trong mỗi phép thử, nếu ngưỡng tin cậy được chọn là giá trị p ≤ 5% để kết luận có sự sai khác có ý nghĩa thống kê thì vì nhiều marker được sử dụng nên sẽ có nhiều phép thử và lúc đó xác suất để 1 phép thử ngẫu nhiên có giá trị p ≤ 5% là rất lớn (dương tính giả - false positive). Do đó cần phải dùng các kỹ thuật thống kê hiệu chỉnh, ví dụ như phương pháp Bonferonni, Benjamini – Hochberg FDR hay hoán vị (permutation) để chọn một ngưỡng giá trị p thích hợp.Trong phân tích QTL bằng phương pháp interval mapping, thường thì với giá trị LOD từ 3 trở lên, khả năng tại vị trí QTL đó có liên quan đến tính trạng cần quan tâm là cao (Hình 2).
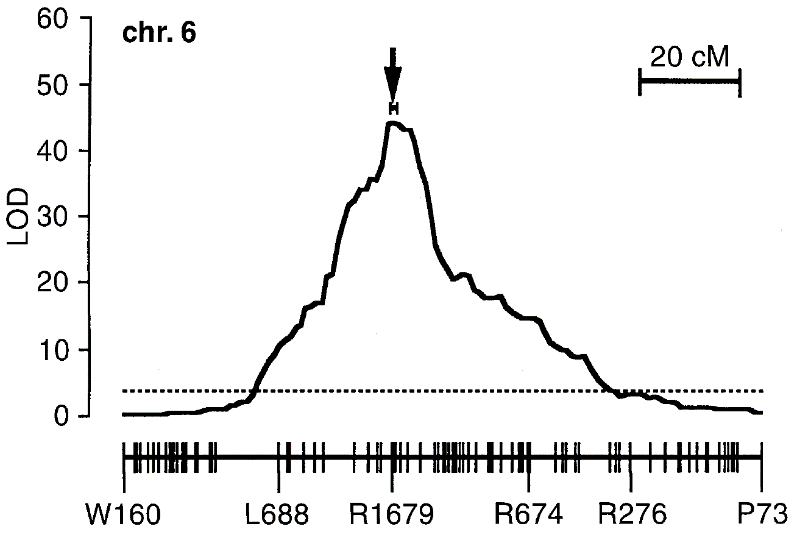 Hình 2. Giá trị LOD được dựng dọc theo chiều dài nhiễm sắc thể với vị trí các marker. Hình thể hiện một đỉnh QTL với giá trị LOD =46. Đường gạch đứt đoạn thể hiện ngưỡng LOD =3 thường được dùng để xem xét sự liên kết giữa marker và tính trạng. Hình được trích dẫn Yano et al (3)
Hình 2. Giá trị LOD được dựng dọc theo chiều dài nhiễm sắc thể với vị trí các marker. Hình thể hiện một đỉnh QTL với giá trị LOD =46. Đường gạch đứt đoạn thể hiện ngưỡng LOD =3 thường được dùng để xem xét sự liên kết giữa marker và tính trạng. Hình được trích dẫn Yano et al (3)
Các tác giả xác định được 5 QTL cho số lượng hạt trên bông và 4 QTL cho chiều cao cây. Đối với tính trạng số lượng hạt trên bông, giá trị LOD dao động từ 1.5-9. Một giá trị nữa trong phân tích QTL cần được chú ý là giá trị R2 hay PVE (Phenotypic variance explained - phần trăm giải thích sự khác biệt kiểu hình tại vị trí QTL đó). Giá trị này có thể tính từ phép phân tích phương sai ANOVA đề cập ở trên. Không ngạc nhiên khi QTL Gn1 có giá trị LOD =9 cũng có giá trị R2 cao nhất: 44%. Cả hai giá trị LOD và R2 cho QTL này là rất lớn, có thể dễ dàng phân biệt được các nhóm kiểu hình khác nhau (QTL này làm tăng số lượng hạt tới 92 hạt). Do đó, xác suất có gene trong vùng QTL này ảnh hưởng đến tính trạng là rất cao (LOD = 9, khả năng có QTL cao hơn 109 lần so với khả năng không có QTL). Đối với tính trạng chiều cao cây, các tác giả cũng xác định được một QTL lớn là Ph1 (LOD = 6.5, R2 = 30%). QTL này nằm trong vùng gene “cách mạng xanh” semi-dwarf1 đã được phân lập và giải trình tự (4). Đọc trình tự cho thấy đúng là Habataki, giống được chọn làm bố trong phép lai này, có đột biến mất đoạn ở gene này. Kết quả này có thể coi như tiêu chuẩn so sánh cho thấy thí nghiệm QTL ở đây là chính xác. Có một điểm thú vị là giống Koshihikari với cây cao và số lượng hạt ít hơn nhưng có 1 QTL tăng chiều cao cây và 3 QTL giảm chiều cao, 4 QTL tăng số lượng hạt trên bông và 2 QTL giảm số lượng hạt. Có thể thấy số QTL làm tăng hay giảm tính trạng kiểu hình không phản ánh kiểu hình tính trạng cuối cùng của cá thể. Trên thực tế, các QTL sẽ tương tác với nhau để tạo ra tính trạng.
Như vậy từ toàn bộ hệ genome với 12 nhiễm sắc thể, bằng kĩ thuật QTL mapping, các tác giả đã khoanh vùng các vị trí có thể có liên quan đến tính trạng cần nghiên cứu. Để tiếp tục tìm ra chính xác gene ảnh hưởng, họ chọn QTL có giá trị lớn nhất để phân lập. Bước tiếp theo này gọi là fine mapping, xác đinh vị trí QTL chính xác đến cụ thể từng gene. Để thuận tiện và tăng chính xác cho quá trình tìm vị trí gene ở độ phân giải cao, các tác giả phát triển dòng đẳng gene (nearly isogenic line -NIL) cho vị trí QTL cần nghiên cứu bằng cách lai lại nhiều lần với một trong hai bố mẹ và sử dụng marker phân tử để chọn lọc. Các dòng gần đẳng gene này giúp tạo sự đồng nhất cho quần thể phân li, giảm hiện tượng tương tác gene ảnh hưởng đến việc đo đạc kiểu hình (Mendel hóa tính trạng số lượng). Các dòng này được dùng làm bố mẹ để tạo mới một quần thể F2. Phân tích nhanh bằng cách sử dụng 96 cây F2 từ dòng đẳng gene NIL-Gn1 các tác giả xác định được trên thực tế có tới 2 QTL Gn1a và Gn1b ở vùng này. QTL Gn1a được xác định chính xác nằm trong khoảng 2 marker R3192 và C12072S, do đó được chọn để tách dòng. Ngoài ra, vùng Gn1a này cũng đã xác định được bản đồ vật lí với các dòng BAC chứa toàn bộ vùng có 2 marker ở trên (hình 2G trongAshikari et al (1)). Với việc trình tự hệ gene lúa đã được giải mã cộng với xác định chính xác khoảng cách vật lí giữa 2 marker giúp việc xác định gene trở nên dễ dàng hơn vì có thể thiết kế được rất nhiều marker và biết được trật tự của chúng trong vùng cần phân tích. Bài toán lúc này sẽ là sử dụng một quần thể phân li lớn với nhiều marker trong vùng phân tích để xác định các cá thể có trao đổi chéo, so sánh với kiểu hình để dần dần thu hẹp độ lớn vùng cần tìm kiếm. Ở đây các tác giả sử dụng tới hơn 13000 cây F2, song cũng cần chú ý là các tác giả không cần xác định kiểu hình trên toàn bộ 13000 cây mà chỉ trên các cây có hiện tượng trao đổi chéo. Khi độ lớn của vùng tìm kiếm càng nhỏ, tỉ lệ trao đổi chéo sẽ rất thấp và cần nhiều cá thể để có xác suất tìm ra cây có trao đổi chéo trong vùng nhỏ đó (1cM tương đương với tỉ lệ 1% trao đổi chéo ở giao tử và 1cM nhiều khi vẫn là 1 vùng lớn có thể chứa tới vài chục gene). Các tác giả đã thu hẹp được vùng có thể liên quan đến tính trạng với độ lớn chỉ là 6.3 kb và chỉ có 1 gene trong khoảng này, gene cytokinin oxidase OsCKX2. Hình 3 tóm lược các bước cơ bản trong xác định và tách dòng QTL.
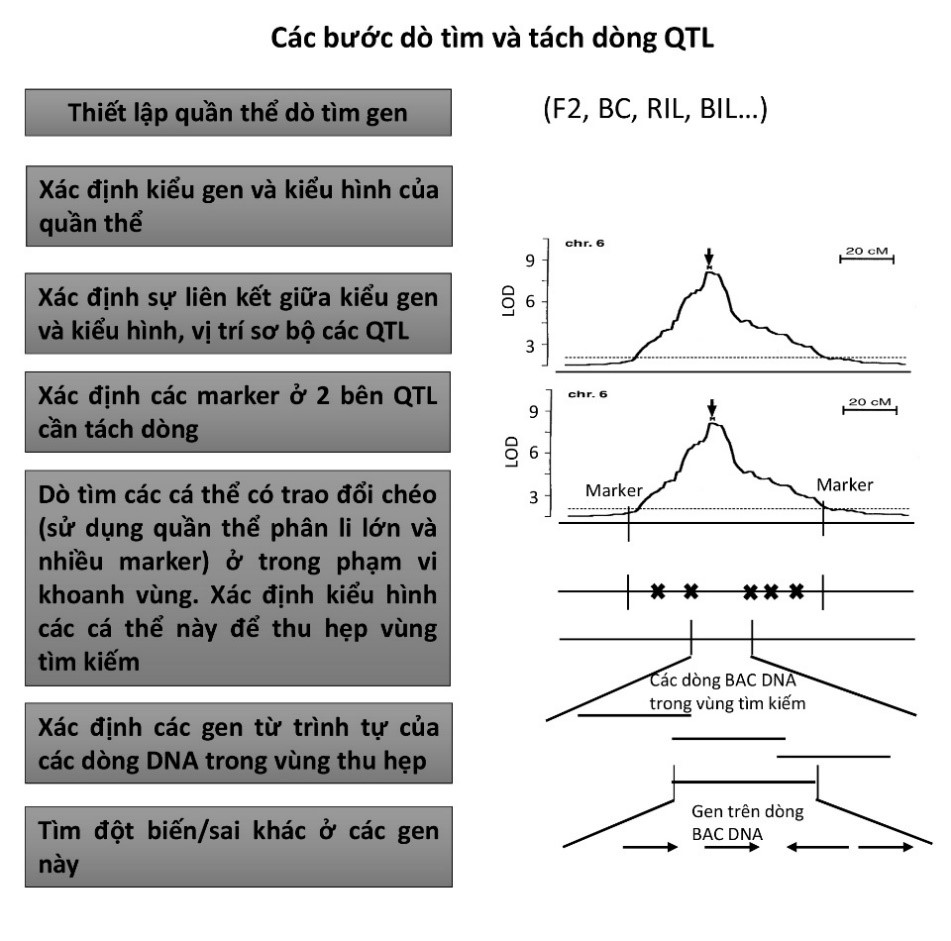 Hình 3. Sơ đồ các bước dò tìm và tách dòng QTL. BC: quần thể lai trở lại. RIL: Recombinant inbred lines - quần thể cận giao tái tổ hợp. BIL: Backcross inbred line - quần thể lai trở lại cận giao tái tổ hợp.
Hình 3. Sơ đồ các bước dò tìm và tách dòng QTL. BC: quần thể lai trở lại. RIL: Recombinant inbred lines - quần thể cận giao tái tổ hợp. BIL: Backcross inbred line - quần thể lai trở lại cận giao tái tổ hợp.
Liệu đã kết luận được là gene này ảnh hưởng đến tính trạng? Để chứng minh điều này đòi hỏi các tác giả cần tìm thêm nhiều allele của gene này và so sánh tính trạng giữa chúng. Nếu có thể các tác giả cũng cần tạo ra các cây chuyển gene làm tăng hay giảm biểu hiện của gene này để xem có thay đổi tương ứng về kiểu hình hay không. Ashikari et al (1) đã tiến hành tìm thêm nhiều giống có các allele khác nhau, đồng thời cũng tạo cây chuyển gene và thể hiện rõ rằng việc tăng hay giảm biểu hiện của gene OsCKX2 ảnh hưởng đến số lượng hạt. Các kết quả này đã chứng minh được sự khác biệt về biểu hiện của gene này chính là bản chất di truyền của QTL đang nghiên cứu. Các tác giả cũng tiến hành thêm 1 bước nữa là áp dụng gene này cho chọn giống, kết hợp cả với allele làm thấp cây và đã cho thấy việc áp dụng vùng nhiễm sắc thể chứa locus Gn1 làm tăng số lượng hạt trên cây tới 30%. Có một điểm cần chú ý ở đây là ở locus Gn1, dạng allele cho năng suất cao xuất phát từ dòng indica, và có thể chỉ có lợi khi chuyển vào dòng japonica. Do đó đối với các nước trồng lúa indica như Việt Nam việc áp dụng QTL này có thể sẽ chưa có nhiều lợi ích.
Kết luận
Tóm lại đây là một nghiên cứu hoàn chỉnh về xác định gene cho một tính trạng khó là năng suất lúa.Với việc thiết kế thí nghiệm chính xác và tiến hành các phương pháp nghiên cứu cẩn thận, công phu đã giúp nhóm nghiên cứu thành công. Bài báo đã thiết lập một tiêu chuẩn cao để các nghiên cứu về di truyền tính trạng số lượng hướng tới.
Đối với Việt Nam, nước ta có nhiều giống cây trồng quý đặc hữu có các tính trạng nông học và chất lượng cao. Việc phân lập gene dựa trên các phương pháp di truyền và sinh học phân tử như ở đây không những nâng cao hiểu biết về các quá trình sinh học ở sinh vật mà còn đóng góp trực tiếp cho việc chọn tạo giống cây trồng có định hướng một cách chính xác. Các tiến bộ kỹ thuật hiện nay đã giúp giảm giá thành việc xác định kiểu gene nhiều lần và là một lợi thế chúng ta cần nắm bắt. Việc quan trọng nhất vẫn sẽ là bảo tồn các nguồn gene quý trong các trung tâm giống cây trồng quốc gia, và thiết kế các thí nghiệm để lai tạo các quần thể và khảo sát kiểu hình, tìm ra gene hay chỉ thị phân tử hữu ích để áp dụng vào sản xuất.
Tài liệu tham khảo:
Ashikari M, et al. (2005) Cytokinin oxidase regulates rice grain production. Science 309(5735):741-745.
Lander ES & Botstein D (1989) Mapping Mendelian factors underlying quantitative traits using RFLPlinkage maps. Genetics 121(1):185-199.
Yano M, et al. (1997) Identification of quantitative trait loci controlling heading date in rice using a high-density linkage map. Theoretical and Applied Genetics 95(7):1025-1032.
Sasaki A, et al. (2002) Green revolution: a mutant gibberellin-synthesis gene in rice. Nature 416(6882):701-702.
Đọc thêm
Tanksley SD (1993) Mapping polygenes. Annual Review of Genetics 27:205-233.
Yano M (2001) Genetic and molecular dissection of naturally occurring variation. Current Opinion in Plant Biology 4(2):130-135.
Lời cảm ơn: Tác giả xin cảm ơn Quỹ Giáo dục Việt Nam VEF đã tài trợ một phần cho nghiên cứu này
Về tác giả: T.S. Nguyễn Viết Cường nhận bằng tiến sĩ về di truyền thực vật tại đại học Cornell, New York, Hoa Kỳ năm 2014. Tác giả nghiên cứu về di truyền cà chua, cụ thể là tìm ra các gene ảnh hưởng đến quá trình phát triển và chín quả và các gene làm tăng chất lượng và giá trị dinh dưỡng ở cà chua.


Add new comment